О НЕВОЗМОЖНОСТИ СЛУЧАЙНОЙ ЭВОЛЮЦИИ И ОБУЧЕНИЯ ОРГАНИЗМОВ
(Екатеринбург)
Рассмотрено поведение организма при регистрации
рецептором нового образа. Показано, что организм не может получать новую ценную
информацию извне (т.е. обучаться). С учетом блочно-иерархической структуры
генома найдена вероятность образования видов за счет случайных мутаций, которая
оказалась исчезающе малой. С точки зрения синтетической теории эволюции (когда
нет программы эволюции) нет оснований говорить о том, что блочно-иерархический
принцип построения генома будет ускорять возникновение новых видов. Необходимо
детерминистское описание эволюции организмов.
ON IMPOSSIBILITY
OF ORGANISMS RANDOM EVOLUTION AND LEARNING
Melkikh A.V.
(Ekaterinburg)
Organism behavior during new pattern
registration is considered. It is shown that organism can’t receive new
valuable information from environment (i.e. to learn). With hierarchical genome
structure the probability of species origin by random mutations is obtained.
This probability was found negligibly small. From the point of view of
synthetic theory of evolution (when the program of evolution is absent) there
is no reason to assume that hierarchical principle of genome structure will
accelerate new species origin. It is necessary to create deterministic
description of organisms.
Предметом рассмотрения данной статьи являются два основных положения биологической науки: синтетическая теория эволюции и возможность обучения организма. Математическое моделирование многих биологических процессах явно или неявно основано на этих положениях. Однако при детальном анализе обоих положений обнаруживаются противоречия.
Рассмотрим их подробнее.
1. СУЩЕСТВУЕТ ЛИ ОБУЧЕНИЕ?
Предмет исследования – информационная система, способная к рецепции информации, ее обработке и принятию решений.
Нас будет интересовать не любая система, а только та, которая получает всю информацию об окружающей среде через рецепторы. Нетрудно видеть, что поведение живых организмов как раз удовлетворяет указанным выше условиям. То есть всю информацию организм получает только через рецепторы.
Что понимается под обучением нейронной сети? Фактически, это задание ее начальных условий, которое не происходит самопроизвольно, а осуществляется человеком извне [1].
Что же происходит в системе, когда она встречается с новым образом, эталона которого не существует в организме?
Проведем мысленный эксперимент. Пусть в системе имеется М эталонов A1,…, AМ. Пусть система встречается с новой ситуацией (новым образом), для которой не существует эталона из имеющегося набора. С одной стороны, система может просто не заметить этот образ и не предпринимать никаких действий (так же как, например, ультрафиолетовый свет не приводит ни к каким действиям при отсутствии соответствующих зрительных рецепторов). С другой стороны, система могла бы попытаться каким-нибудь образом обработать этот сигнал. Например, сделать его новым эталоном. Но как узнать, годится ли новый образ для этого? Допустим, что в системе существует алгоритм формирования новых эталонов. Этот алгоритм может быть реализован только на основе уже имеющихся эталонов:
![]() .
.
Однако любой алгоритм работает лишь с определенным набором параметров. Следовательно, прежде чем такой алгоритм запустится, необходимо распознать образ (сделать вывод о том, к какому классу объектов он относится). Кроме того, AM+1 не может быть эталоном, поскольку основным свойством эталонов является их независимость друг от друга. Такое свойство имеет место, например, в алфавите, используемом машиной Тьюринга.
Что нам делать с этим образом? Сделать его новым эталоном? А если это совершенно бесполезная информация?
Самое главное заключается в следующем: сделать образ новым эталоном – не значит просто записать в память! Каждому эталону должна соответствовать программа, сопоставляющая этому эталону определенное действие эффектора. Эталон без программы совершенно бесполезен, поскольку в этом случае на работе эффекторов его существование никак не скажется (поведение организма не изменится). В окружающей среде, однако, нет никаких программ, а только образы. Следовательно, программы работы с новыми эталонами получить из окружающей среды невозможно. Если их нет, то они и не появятся.
Если попытаться использовать новый эталон и старые программы, то это можно сделать единственным способом (поскольку старая программа «не понимает» новый эталон) – идентифицировать этот новый эталон с одним из старых эталонов. Однако при этом никакого нового действия не произойдет. Организм при этом просто путает этот новый образ с одним из старых. При этом его действия будут неадекватны. Организм этого даже не поймет, и будет действовать так и дальше, пока не погибнет от непроизводительных затрат энергии! Естественно, что он при этом ничему не научится.
Почему же тогда при наблюдении за организмом, мы видим, что эффекторы запускаются не сразу, а только после неоднократного предъявление сигнала (именно это мы и называем обучением)?
Наиболее выгодной для организма может оказаться следующая стратегия:
провести серию измерений, при этом не запускать эффектор (это может быть не
выгодно), после этого найти среднее значение величины (уменьшая тем самым
случайную погрешность), и только потом запустить эффектор, соответствующий
этому среднему образу.
То есть эффектор не запускается сразу не потому, что нет программы действия, и она формируется, а потому, что выгодно провести серию измерений и лишь потом начать действовать. В противном случае организм может проиграть в энергетических затратах, и, как следствие – погибнуть.
Вывод: обучающихся
интеллектуальных систем не существует. Все действия организмов (а так же
искусственных систем) могут и должны быть описаны на основе идеологии
адаптирующихся автоматов, которые не могут обучаться, а лишь выполняют
априорные программы.
2. МОГЛА ЛИ
ЖИЗНЬ ЭВОЛЮЦИОНИРОВАТЬ ПУТЕМ СЛУЧАЙНЫХ МУТАЦИЙ?
В современной научной литературе существуют две точки зрения на эволюцию видов. Первая из них основана на классических представлениях синтетической теории эволюции [3] и сводится к тому, что новые виды образуются за счет случайных мутаций под действием естественного отбора. Вторая точка зрения связана с различными представлениями о детерминированности эволюционных процессов [4].
Одним из критериев правильности эволюционных теорий является, например, их способность описать экспериментальные данные по скорости возникновения новых видов.
Проблеме скорости возникновения видов посвящено множество работ, как теоретических, так и экспериментальных. В частности, в ряде работ [5-8] рассматривается блочно-иерархический принцип организации генома. То есть, если геном иерархически структурирован, то возможных сочетаний в нем гораздо меньше, чем для случая независимых нуклеотидов. Проведенные оценки для простейших организмов показывают, что блочные мутации приводят к вполне разумным временам возникновения новых видов.
Однако неясным остается ключевой вопрос, какая система будет делить ДНК на блоки, и определять их состав? Согласуется ли представление о блочных мутациях с дарвиновским механизмом образования новых видов?
Оценим вероятность возникновения видов за счет случайных мутаций.
Независимые нуклеотиды. Будем считать сначала, что мутации
различных нуклеотидов независимы друг от друга и имеют характер случайного
процесса (в соответствии с теорией Дарвина). Первые же оценки показывают, что
число возможных комбинаций нуклеотидов (без учета изменения общего их числа)
составит порядка ![]() (поскольку нуклеотидов четыре вида, а число нуклеотидов в ДНК
высших организмов примерно
(поскольку нуклеотидов четыре вида, а число нуклеотидов в ДНК
высших организмов примерно ![]() ). Это число значительно больше, чем количество всех организмов,
когда-либо живших на Земле. Такие оценки делались и раньше [6-8], однако в них
не учитывалось реальное генетическое различие между видами.
). Это число значительно больше, чем количество всех организмов,
когда-либо живших на Земле. Такие оценки делались и раньше [6-8], однако в них
не учитывалось реальное генетическое различие между видами.
Пусть m – количество организмов одного вида, t – характерное время возникновения нового вида, T – время жизни одного организма, k – количество экологических ниш, окружающих данную нишу (потенциально возможные близкие виды), W – вероятность мутации конкретного нуклеотида за время жизни организма. Далее, пусть N – общее число нуклеотидов в ДНК (для упрощения будем считать, что общее число нуклеотидов не меняется, что не повлияет на общность выводов), N1 - число нуклеотидов в ДНК близкого потенциального вида, отличных от нуклеотидов рассматриваемого вида.
Пусть N1 - число нуклеотидов, которые могут изменяться, и при этом организм будет оставаться в пределах одного вида. То есть это характерная ширина «потенциальной ямы» одного вида.
Тогда можно определить число организмов (или вероятность – P), которые путем случайных мутаций доберутся до следующей ниши, то есть образуют новый вид с вполне определенными N1 нуклеотидами.
Учитывая сказанное выше, получим формулу:
![]() .
.
Оценим величину P сверху. Для этого выберем такое значение W, которое соответствует максимуму величины P. На самом деле эта величина зависит от многих внешних факторов (температуры, радиации и т.д.) и может быть далека от оптимальной.
Для нахождения максимума приравняем нулю производную
![]() ,
,
и получим:
![]() и
и ![]() . (1)
. (1)
Найденная величина W очень велика. Действительно, она соответствует ситуации, когда уже в следующем поколении генетические отличия потомков от родителей становятся столь велики, как между отдельными видами. То есть, если мы примем величину W близкой к действительности, то и значение P будет гораздо меньше.
Примем, что k » 10.
Характерное время возникновения вида составляет »![]() лет, среднее время жизни организма »
лет, среднее время жизни организма »![]() лет, а величину m
примем равной
лет, а величину m
примем равной ![]() (для мелких животных
эта величина будет конечно больше, но, как будет видно из оценок, основной
вывод от этого не изменится).
(для мелких животных
эта величина будет конечно больше, но, как будет видно из оценок, основной
вывод от этого не изменится).
Таким образом, получим: mt/T
»
![]() . Величина N у
высших организмов составляет »
. Величина N у
высших организмов составляет »![]() (у простейших меньше, однако этот факт так же не изменит
общего вывода). Используя данные сравнительной биохимии, оценим величины N1 и N2: N1
»
0.01N, N2 = 0.001N.
(у простейших меньше, однако этот факт так же не изменит
общего вывода). Используя данные сравнительной биохимии, оценим величины N1 и N2: N1
»
0.01N, N2 = 0.001N.
Таким образом, окончательно получим:
![]() .
.
Такое число является исчезающе малым, что полностью согласуется с проведенными ранее оценками. То есть для случая независимых нуклеотидов можно сделать вывод о том, что за счет случайных мутаций виды возникнуть не могли. Нетрудно показать, что полученная формула обладает большим запасом прочности (например, количество организмов, а так же различие между видами можно варьировать в широких пределах). Полученный вывод не изменится, если рассматривать самые распространенные и короткоживущие виды вплоть до бактерий.
Блочно-иерархическая структура генома. Теперь рассмотрим важный случай, когда нуклеотиды не ведут себя независимо, а организуются в некие структуры (гены), которые могут претерпевать изменения в процессе эволюции (регуляция генома [9], работа молекулярно-генетических систем управления [5]). Это значительно уменьшает число возможных состояний (процессы самоорганизации).
Рассмотрим, с одной стороны пространство нуклеотидов, а с другой стороны – пространство признаков организмов (рисунок).
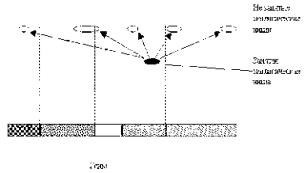
Рисунок
В пространстве нуклеотидов конфигурации генов, хромосом и других структур будут определяться силами взаимодействия молекул. Понятно, что при изменении состава или положения нуклеотидов энергетически выгодными могут оказаться те или иные конфигурации.
В пространстве признаков организмов ситуация иная. Это пространство так же структурировано (в нем существуют экологические ниши, занятые определенными видами), однако положение экологических ниш будет определяться уже приспособленностью всего организма. То есть тем, какие органы он имеет (и сколь они эффективны) для добывания пищи, размножения и т.д.
Но если эволюция происходит за счет случайных мутаций, и у организма нет никакой программы эволюции на будущее (согласно синтетической теории эволюции), то нет никакой связи между структурами нуклеотидов и положением экологических ниш! Легко представить себе, что термодинамически выгодное состояние макромолекулы (ДНК) не только не будет соответствовать организму, способному выжить и оставить потомство, но и вообще никакому организму не будет соответствовать!
Поэтому в случае дарвиновской эволюции организм вынужден будет действовать «в слепую», при этом для попадания в новую нишу популяция должна будет перебрать все возможные комбинации нуклеотидов в каждом гене (выгодность химических структур не обязана соответствовать новой экологической нише, положение которой организму не известно)! Следовательно, формула (1) сохраняет свою силу.
Список литературы:
1.Круглов В.В., Борисов В.В. Искусственные нейронные сети. Теория и практика. М.: Горячая линия “Телеком”, 2001. 361с.
2.Цетлин М.Л. Исследования по теории автоматов и моделирование биологических систем. М.: Наука. 1969. 316с.
3.Воронцов Н.Н. Развитие эволюционных идей в биологии, М.: Издат. отдел УНЦ ДО МГУ, Прогресс-Традиция, АБФ, 1999. 640 с.
4.Берг Л.С. Номогенез или эволюция на основе закономерностей, Петербург, Государственное издательство, 1922. 306 с.
5.Ратнер В.А. Блочно-модульный принцип организации и эволюции молекулярно-генетических систем управления (МГСУ) // Генетика. Т. 28. № 2. С.5-23.
6.Шноль С.Э. Хватает ли времени для дарвиновской эволюции? // Природа. № 11. 1990. С.23-26.
7.Чернавский Д.С. Проблема происхождения жизни и мышления с точки зрения современной физики // Успехи физических наук. Т.170. №2. 2000. С.157-183.
8.Моносов Я.А. О факторах ускорения биологической эволюции // Биофизика. Т. 36, вып.5. 1991. С. 920-922.
9.Singer M., Berg P. Genes and Genomes. University science books, California. 1991.